Строение бактерий. Значение пилей в жизни бактерий Пили характерны для
Наряду со жгутиками прокариоты могут обладать и другими внеклеточными образованиями. В середине ХХ века было установлено, что бактерии способны формировать специфическую группу поверхностных образований. Их называли ворсинками, ресничками, фимбриями. Сегодня их называют пилями бактерий.
Внешне пили, или фимбрии, как их называли до 1956 г., выглядят как микроскопические волоски, покрывающие клетку бактерии. На 1 клетку прокариота может приходиться от нескольких единиц до тысяч ворсинок.
Хотя они, как и жгутики, являются поверхностными образованиями, однако между собой имеют больше различий, чем сходств.
По размеру пили намного меньше жгутиков, в среднем в 3 раза тоньше (не более 10 нм), и длиной не превышают 1,5 мкм.
По строению, несмотря на то, что как пили, так и жгутики состоят из белковых клеток, они также различаются:
- пили, или фимбрии, представляют собой легкую цепочку проводящих белков цилиндрической формы, отходящую от поверхностного слоя клетки;
- жгутики являются более громоздкими по строению, с наличием сложных структур (стержень, базальное тело, кольца и другое).
 Столь явное различие в строении поверхностных образований прокариотов связано с совершенно разными задачами, которые они решают в процессе жизнедеятельности бактериальной клетки.
Столь явное различие в строении поверхностных образований прокариотов связано с совершенно разными задачами, которые они решают в процессе жизнедеятельности бактериальной клетки.
Для чего прокариотам пили
К примеру, если жгутики бактерий обеспечивают возможность передвигаться, то фимбрии не имеют никакого отношения к перемещению в пространстве и присутствуют как у движущихся, так и у неподвижных бактерий.
В отличие от жгутиков функции пилей бактерий изучены достаточно слабо, но совершенно очевидно, что одной из них является способность обеспечить прикрепление клетки бактерии к питательному субстрату.
Разные типы ворсинок
Пили не являются однородными образованиями, их различают как минимум 4 типа, каждый из которых выполняет свои функции, причем одна клетка может являться носителем несколько разных типов фимбрий.
Пили 1 типа
Фимбрии бактерий 1 типа образуются из пилина (белок) и отличаются крайне прочной связью с прокариотом. Чтобы отделить такую фимбрию от бактериальной клетки, требуются усилия, многократно превышающие необходимое воздействие для отделения половых пилей или жгутиков.

Для пилей 1 типа характерно расположение перитрихиально – по всей поверхности бактерии.
Исследования методами выявления свойств показали, что пили 1 типа являются химически устойчивыми образованиями – они инертны к растворам щелочей, мочевине и трипсину (фермент, расщепляющий белки).
Разрушаются пили 1 типа при кипячении в растворах с высокой кислотностью, при этом методе воздействия происходит необратимое разрушение (денатурация) белка, образующего фимбрию.
Характерной особенностью пилей 1 типа является:
- способность образовывать пленки и придавать бактериям гидрофобные свойства;
- способность вызывать агглютинацию эритроцитов (выпадение в осадок в результате склеивания) под действием агглютининов.

Органоиды бактерий
Основными функциями являются:
- адгезивная – прикрепление бактерий к субстратам;
- защитная – объединение клеток прокариотов, получивших гидрофобные свойства, в группы;
- участие в процессах метаболизма клетки – увеличение всасывающей поверхности.
Ворсинки 2 типа
Эта группа имеет очень много общего с предыдущей, однако не обладает характерными особенностями 1 типа – пили не участвуют в формировании пленок и не приклеиваются к эритроцитам (агглютинация), провоцируя выпадение их в осадок.
Столь близкое сходство позволяет предположить, что пили 2 типа являются мутантной формой 1 типа.
Половые фимбрии (3 тип)
Современные методы выявления позволили определить, что при горизонтальном переносе генетического материала (конъюгации) ключевую роль играют половые пили.
Возможность непосредственного контакта двух бактериальных клеток с последующей конъюгацией была выявлена в 50-х годах прошлого века в результате исследований двух американских биохимиков – Д.Ледербейга и Э.Тейтема. Данный процесс имеет большое практическое значение, так как позволяет производить обмен наследственными признаками организмам, размножающимся только прямым делением.
Половые фимбрии, их называют F-пили, присутствуют только у бактериальных штаммов, обладающих фактором трансмиссивности – это может быть автономный репликон или его часть.
F-пили представляют собой цилиндрические белковые образования большего диаметра, чем пили 1 или 2 типа, расположенные перпендикулярно к поверхности.
 Формирование пиля осуществляется на поверхности цитоплазматической мембраны в точках контакта ее с вешней оболочкой. Сформированная трубочка проходит сквозь слои муреина и внешнюю мембрану.
Формирование пиля осуществляется на поверхности цитоплазматической мембраны в точках контакта ее с вешней оболочкой. Сформированная трубочка проходит сквозь слои муреина и внешнюю мембрану.
В случае потери F-пили восстанавливаются – в течение 30 секунд пиль достигает половины своей величины. Для формирования полноценной трубочки необходимо от 4 до 5 минут.
Современные методы выявления позволили определить, что на поверхности бактерии F-пили сохраняются в течение 5 минут, после чего сбрасываются, и процесс повторяется.
F-пили значительно отличаются от ворсинок 1 и 2 типа как по строению, так и по свойствам.
В отличие от последних F-пили легко отделяются от бактериальной клетки даже при незначительном встряхивании.
Методами физико-химического анализа было определено, что в составе F-пили отсутствует целый ряд α-аминокислот, характерных для белка пилей 1 типа, но через ковалентную связь присоединены остаток D-глюкозы и две фосфатные группы.
В связи с иной химической структурой на F-пили не адсорбируются обычные фаги, а только специфичные для них, называемые мужскими фагами.
Участие F-пили в процессе передачи информации
Процесс передачи части генетической информации подразумевает наличие пары клетка-донор и клетка-реципиент.
- Первоначально клетка-донор формирует F-пиль.
- F-пиль донора фиксируется на клетке-реципиенте.
- В F-плазмиде донорной клетки осуществляется разрыв одной нити ДНК, которая передается реципиенту.
- Обе бактерии достраивают вторую цепочку ДНК и восстанавливают F- плазмиду. Клетка-реципиент превращается в донора.

Микроскопические методы исследований позволили определить, что образование F-пилей характерно только для растущих и активных клеток, при переходе в стационарную фазу роста бактерии теряют свою способность образовывать половые пили и становятся плохими донорами.
Специфическая направленность фимбрий 4 типа
Пили 4 типа принимают участие в обеспечении разновидности скользящего движения бактерий всей колонией.
 Сам процесс скольжения с участием пилей 4 группы предполагает наличие 2 систем движения:
Сам процесс скольжения с участием пилей 4 группы предполагает наличие 2 систем движения:
- А-система — секретирует слизь на полюсе направления движения микроорганизма.
- S-система – роение; обеспечивается последовательным сокращением и удлинением пилей 4 типа, подобное подтягиванию.
Механизм данного вида бактериального движения на сегодняшний день находится в процессе изучения, и большинство выводов носят предположительный характер.
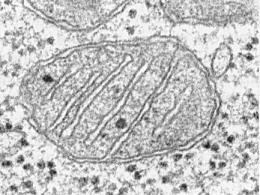
К поверхностным структурам бактериальной клетки относятся также ворсинки (фимбрии, пили) ( рис. 4 , 6). Их насчитывается от нескольких единиц до нескольких тысяч на клетку. Эти структуры не имеют отношения к движению бактерий и обнаружены у подвижных и неподвижных форм. Ворсинки построены из одного вида белка - пилина - и представляют собой прямые белковые цилиндры, отходящие от поверхности клетки. Они, как правило, тоньше жгутиков (диаметр - 5-10 нм, длина 0,2-2,0 мкм), расположены перитрихиально или полярно. Больше всего сведений имеется о ворсинках Е. coli . У этой бактерии описаны ворсинки общего типа и половые.
Ворсинки общего типа придают бактериям свойство гидрофобности, обеспечивают их прикрепление к клеткам растений, грибов и неорганическим частицам, принимают участие в транспорте метаболитов. Через ворсинки в клетку могут проникать вирусы .
Наиболее хорошо изучены половые ворсинки, или F-пили, принимающие участие в половом процессе бактерий. F-пили необходимы клетке-донору для обеспечения контакта между ней и реципиентом и в качестве конъюгационного тоннеля, по которому происходит передача ДНК. Ворсинки нельзя считать обязательной клеточной структурой, так как и без них бактерии хорошо растут и размножаются.
Фимбрии (пили) - нитевидные белковые органеллы, покрывающих всю поверхность бактериальной клетки - антигены фактора колонизации . Эти тонкие структуры позволяют бактерии прикрепляться к эпителиальным клеткам и препятствуют ее захвату нейтрофилами
Фимбрии состоят из множества одинаковых белковых субъединиц. Эта субъединица называется пилином (молекулярная масса 17000-30000). В составе пилина есть консервативные и вариабельные участки. Перестройки хромосом, ведущие к экспрессии любого из множества неактивных генов пилина, сопровождаются изменениями антигенного состава фимбрий.
При электронной микроскопии фимбрии выглядят как похожие на волоски выросты, проникающие через наружную мембрану. Они могут располагаться на одном конце клетки либо более равномерно по всей ее поверхности. У отдельной клетки может быть несколько сотен фимбрий, которые выполняют различные функции.
У некоторых фимбрий (например, у дигалактозидсвязывающих фимбрий Escherichia coli) на апикальном конце находятся специальные белки, играющие важную роль во взаимодействии с рецепторами клеток.
Считается, что главная функция фимбрий - обеспечение фиксации бактерий в тканях.
Структура бактерий хорошо изучена с помощью электронной микроскопии целых клеток и их ультратонких срезов, а также других методов. Бактериальную клетку окружает оболочка, состоящая из клеточной стенки и цитоплазматической мембраны. Под оболочкой находится протоплазма, состоящая из цитоплазмы с включениями и наследственного аппарата - аналога ядра, называемого нуклеоидом (рис. 2.2). Имеются дополнительные структуры: капсула, микрокапсула, слизь, жгутики, пили. Некоторые бактерии в неблагоприятных условиях способны образовывать споры.
Рис. 2.2. Структура бактериальной клетки: 1 - капсула; 2 - клеточная стенка; 3 - цитоплазматическая мембрана; 4 - мезосомы; 5 - нуклеоид; 6 - плазмида; 7 - рибосомы; 8 - включения; 9 - жгутик; 10 - пили (ворсинки)
Клеточная стенка - прочная, упругая структура, придающая бактерии определенную форму и вместе с подлежащей цитоплазматической мембраной сдерживающая высокое осмотическое давление в бактериальной клетке. Она участвует в процессе деления клетки и транспорте метаболитов, имеет рецепторы для бактериофагов, бактериоцинов и различных веществ. Наиболее толстая клеточная стенка у грамположительных бактерий (рис. 2.3). Так, если толщина клеточной стенки грамотрицательных бактерий около 15-20 нм, то у грамположительных она может достигать 50 нм и более.
Основу клеточной стенки бактерий составляет пептидогликан. Пептидогликан является полимером. Он представлен параллельными полисахаридными гликановыми цепями, состоящими из повторяющихся остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных гликозидной связью. Эту связь разрывает лизоцим, являющийся ацетилмурамидазой.
К N-ацетилмурамовой кислоте ковалентными связями присоединен тетрапептид. Тетрапептид состоит из L-аланина, который связан с N-ацетилмурамовой кислотой; D-глутамина, который у грамположительных бактерий соединен с L-лизином, а у грамотри-
 Рис. 2.3.
Схема архитектоники клеточной стенки бактерий
Рис. 2.3.
Схема архитектоники клеточной стенки бактерий
цательных бактерий - с диаминопимелиновой кислотой (ДАП), которая представляет собой предшественник лизина в процессе бактериального биосинтеза аминокислот и является уникальным соединением, присутствующим только у бактерий; 4-й аминокислотой является D-аланин (рис. 2.4).
В клеточной стенке грамположительных бактерий содержится небольшое количество полисахаридов, липидов и белков. Основным компонентом клеточной стенки этих бактерий является многослойный пептидогликан (муреин, мукопептид), составляющий 40-90% массы клеточной стенки. Тетрапептиды разных слоев пептидогликана у грамположительных бактерий соединены друг с другом полипептидными цепочками из 5 остатков глицина (пентаглицина), что придает пептидогликану жесткую геометрическую структуру (рис. 2.4, б). С пептидогликаном ктеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. tekhos - стенка), молекулы которых представляют собой цепи из 8-50 остатков глицерола и рибитола, соединенных фосфатными мостиками. Форму и прочность бактериям придает жесткая волокнистая структура многослойного, с поперечными пептидными сшивками пептидогликана.
 Рис. 2.4.
Структура пептидогликана: а - грамотрицательные бактерии; б - грамположительные бактерии
Рис. 2.4.
Структура пептидогликана: а - грамотрицательные бактерии; б - грамположительные бактерии
Способность грамположительных бактерий при окраске по Граму удерживать генциановый фиолетовый в комплексе с йодом (сине-фиолетовая окраска бактерий) связана со свойством многослойного пептидогликана взаимодействовать с красителем. Кроме этого последующая обработка мазка бактерий спиртом вызывает сужение пор в пептидогликане и тем самым задерживает краситель в клеточной стенке.
Грамотрицательные бактерии после воздействия спиртом утрачивают краситель, что обусловлено меньшим количеством пептидогликана (5-10% массы клеточной стенки); они обесцвечиваются спиртом, и при обработке фуксином или сафранином приобретают красный цвет. Это связано с особенностями строения клеточной стенки. Пептидогликан в клеточной стенке грамотрицательных бактерий представлен 1-2 слоями. Тетрапептиды слоев соединены между собой прямой пептидной связью между аминогруппой ДАП одного тетрапептида и карбоксильной группой D-аланина тетрапептида другого слоя (рис. 2.4, а). Кнаружи от пептидогликана расположен слой липопротеина, соединенный с пептидогликаном через ДАП. За ним следуетнаружная мембрана клеточной стенки.
Наружная мембрана является мозаичной структурой, представленной липополисахаридами (ЛПС), фосфолипидами и белками. Внутренний слой ее представлен фосфолипидами, а в наружном слое расположен ЛПС (рис. 2.5). Таким образом, наружная мем-
 Рис. 2.5.
Структура липополисахарида
Рис. 2.5.
Структура липополисахарида
брана асимметрична. ЛПС наружной мембраны состоит из трех фрагментов:
Липида А - консервативной структуры, практически одинаковой у грамотрицательных бактерий. Липид А состоит из фосфорилированных глюкозоаминовых дисахаридных единиц, к которым прикреплены длинные цепочки жирных кислот (см. рис. 2.5);
Ядра, или стержневой, коровой части (от лат. core - ядро), относительно консервативной олигосахаридной структуры;
Высоковариабельной О-специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями.
ЛПС заякорен в наружной мембране липидом А, обусловливающим токсичность ЛПС и отождествляемым поэтому с эндотоксином. Разрушение бактерий антибиотиками приводит к освобождению большого количества эндотоксина, что может вызвать у больного эндотоксический шок. От липида А отходит ядро, или стержневая часть ЛПС. Наиболее постоянной частью ядра ЛПС является кетодезоксиоктоновая кислота. О-специфическая полисахаридная цепь, отходящая от стержневой части молекулы ЛПС,
состоящая из повторяющихся олигосахаридных единиц, обусловливает серогруппу, серовар (разновидность бактерий, выявляемая с помощью иммунной сыворотки) определенного штамма бактерий. Таким образом, с понятием ЛПС связаны представления об О-антигене, по которому можно дифференцировать бактерии. Генетические изменения могут привести к дефектам, укорочению ЛПС бактерий и появлению в результате этого шероховатых колоний R-форм, теряющих О-антигенную специфичность.
Не все грамотрицательные бактерии имеют полноценную О-специфическую полисахаридную цепь, состоящую из повторяющихся олигосахаридных единиц. В частности, бактерии рода Neisseria имеют короткий гликолипид, который называется липоолигосахаридом (ЛОС). Он сравним с R-формой, потерявшей О-антигенную специфичность, наблюдаемой у мутантных шероховатых штаммов E. coli. Структура ЛОС напоминает структуру гликосфинголипида цитоплазматической мембраны человека, поэтому ЛОС мимикрирует микроб, позволяя ему избегать иммунного ответа хозяина.
Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемыепоринами, окаймляют гидрофильные поры, через которые проходят вода и мелкие гидрофильные молекулы с относительной массой до 700 Д.
Между наружной и цитоплазматической мембраной находится периплазматическое пространство, или периплазма, содержащая ферменты (протеазы, липазы, фосфатазы, нуклеазы, β-лактамазы), а также компоненты транспортных систем.
При нарушении синтеза клеточной стенки бактерий под влиянием лизоцима, пенициллина, защитных факторов организма и других соединений образуются клетки с измененной (часто шаровидной) формой: протопласты - бактерии, полностью лишенные клеточной стенки; сферопласты - бактерии с частично сохранившейся клеточной стенкой. После удаления ингибитора клеточной стенки такие измененные бактерии могут реверсировать, т.е. приобретать полноценную клеточную стенку и восстанавливать исходную форму.
Бактерии сфероили протопластного типа, утратившие способность к синтезу пептидогликана под влиянием антибиотиков или других факторов и способные размножаться, называются L-формами (от названия Института им. Д. Листера, где они впер-
вые были изучены). L-формы могут возникать и в результате мутаций. Они представляют собой осмотически чувствительные, шаровидные, колбовидные клетки различной величины, в том числе и проходящие через бактериальные фильтры. Некоторые L-формы (нестабильные) при удалении фактора, приведшего к изменениям бактерий, могут реверсировать, возвращаясь в исходную бактериальную клетку. L-формы могут образовывать многие возбудители инфекционных болезней.
Цитоплазматическая мембрана при электронной микроскопии ультратонких срезов представляет собой трехслойную мембрану (2 темных слоя толщиной по 2,5 нм каждый разделены светлым - промежуточным). По структуре она похожа на плазмолемму клеток животных и состоит из двойного слоя липидов, главным образом фосфолипидов, с внедренными поверхностными, а также интегральными белками, как бы пронизывающими насквозь структуру мембраны. Некоторые из них являются пермеазами, участвующими в транспорте веществ. В отличие от эукариотических клеток, в цитоплазматической мембране бактериальной клетки отсутствуют стеролы (за исключением микоплазм).
Цитоплазматическая мембрана является динамической структурой с подвижными компонентами, поэтому ее представляют как мобильную текучую структуру. Она окружает наружную часть цитоплазмы бактерий и участвует в регуляции осмотического давления, транспорте веществ и энергетическом метаболизме клетки (за счет ферментов цепи переноса электронов, аденозинтрифосфатазы - АТФазы и др.). При избыточном росте (по сравнению с ростом клеточной стенки) цитоплазматическая мембрана образует инвагинаты - впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Менее сложно закрученные структуры называются внутрицитоплазматическими мембранами. Роль мезосом и внутрицитоплазматических мембран до конца не выяснена. Предполагают даже, что они являются артефактом, возникающим после приготовления (фиксации) препарата для электронной микроскопии. Тем не менее считают, что производные цитоплазматической мембраны участвуют в делении клетки, обеспечивая энергией синтез клеточной стенки, принимают участие в секреции веществ, спорообразовании, т.е. в процессах с высокой затратой энергии. Цитоплазма занимает основной объем бактери-
альной клетки и состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул - рибосом, ответственных за синтез (трансляцию) белков.
Рибосомы бактерий имеют размер около 20 нм и коэффициент седиментации 70S, в отличие от 80S-рибосом, характерных для эукариотических клеток. Поэтому некоторые антибиотики, связываясь с рибосомами бактерий, подавляют синтез бактериального белка, не влияя на синтез белка эукариотических клеток. Рибосомы бактерий могут диссоциировать на две субъединицы: 50S и 30S. рРНК - консервативные элементы бактерий («молекулярные часы» эволюции). 16S-рРНК входит в состав малой субъединицы рибосом, а 23S-рРНК - в состав большой субъединицы рибосом. Изучение 16S рРНК является основой геносистематики, позволяя оценить степень родства организмов.
В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, β-оксимасляной кислоты и полифосфатов (волютин). Они накапливаются при избытке питательных веществ в окружающей среде и выполняют роль запасных веществ для питания и энергетических потребностей.
Волютин обладает сродством к основным красителям и легко выявляется с помощью специальных методов окраски (например, по Нейссеру) в виде метахроматических гранул. Толуидиновым синим или метиленовым голубым волютин окрашивается в краснофиолетовый цвет, а цитоплазма бактерии - в синий. Характерное расположение гранул волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки. Метахроматическое окрашивание волютина связано с высоким содержанием полимеризованного неорганического полифосфата. При электронной микроскопии они имеют вид электронноплотных гранул размером 0,1-1 мкм.
Нуклеоид - эквивалент ядра у бактерий. Он расположен в центральной зоне бактерий в виде двунитевой ДНК, плотно уложенной наподобие клубка. Нуклеоид бактерий, в отличие от эукариот, не имеет ядерной оболочки, ядрышка и основных белков (гистонов). У большинства бактерий содержится одна хромосома, представленная замкнутой в кольцо молекулой ДНК. Но у некоторых бактерий имеются две хромосомы кольцевой формы (V. cholerae) и линейные хромосомы (см. раздел 5.1.1). Нуклеоид выявляется в световом микроскопе после окраски специфическими для ДНК
методами: по Фельгену или по Романовскому-Гимзе. На электронограммах ультратонких срезов бактерий нуклеоид имеет вид светлых зон с фибриллярными, нитевидными структурами ДНК, связанной определенными участками с цитоплазматической мембраной или мезосомой, участвующими в репликации хромосомы.
Кроме нуклеоида, в бактериальной клетке имеются внехромосомные факторы наследственности - плазмиды (см. раздел 5.1.2), представляющие собой ковалентно замкнутые кольца ДНК.
Капсула, микрокапсула, слизь. Капсула - слизистая структура толщиной более 0,2 мкм, прочно связанная с клеточной стенкой бактерий и имеющая четко очерченные внешние границы. Капсула различима в мазках-отпечатках из патологического материала. В чистых культурах бактерий капсула образуется реже. Она выявляется при специальных методах окраски мазка по Бурри- Гинсу, создающих негативное контрастирование веществ капсулы: тушь создает темный фон вокруг капсулы. Капсула состоит из полисахаридов (экзополисахаридов), иногда из полипептидов, например у сибиреязвенной бациллы она состоит из полимеров D-глутаминовой кислоты. Капсула гидрофильна, включает большое количество воды. Она препятствует фагоцитозу бактерий. Капсула антигенна: антитела к капсуле вызывают ее увеличение (реакция набухания капсулы).
Многие бактерии образуют микрокапсулу - слизистое образование толщиной менее 0,2 мкм, выявляемое лишь при электронной микроскопии.
От капсулы следует отличать слизь - мукоидные экзополисахариды, не имеющие четких внешних границ. Слизь растворима в воде.
Мукоидные экзополисахариды характерны для мукоидных штаммов синегнойной палочки, часто встречающихся в мокроте больных кистозным фиброзом. Бактериальные экзополисахариды участвуют в адгезии (прилипании к субстратам); их еще называют гликокаликсом.
Капсула и слизь предохраняют бактерии от повреждений, высыхания, так как, являясь гидрофильными, хорошо связывают воду, препятствуют действию защитных факторов макроорганизма и бактериофагов.
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие на-
чало от цитоплазматической мембраны, имеют большую длину, чем сама клетка. Толщина жгутиков 12-20 нм, длина 3-15 мкм. Они состоят из трех частей: спиралевидной нити, крюка и базального тельца, содержащего стержень со специальными дисками (одна пара дисков у грамположительных и две пары у грамотрицательных бактерий). Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем - ротором, вращающим жгутик. В качестве источника энергии используется разность протонных потенциалов на цитоплазматической мембране. Механизм вращения обеспечивает протонная АТФ-синтетаза. Скорость вращения жгутика может достигать 100 об/с. При наличии у бактерии нескольких жгутиков они начинают синхронно вращаться, сплетаясь в единый пучок, образующий своеобразный пропеллер.
Жгутики состоят из белка - флагеллина (flagellum - жгутик), являющегося антигеном - так называемый Н-антиген. Субъединицы флагеллина закручены в виде спирали.
Число жгутиков у бактерий разных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен, отходящих по периметру бактерии (перитрих), у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.
Жгутики выявляют с помощью электронной микроскопии препаратов, напыленных тяжелыми металлами, или в световом микроскопе после обработки специальными методами, основанными на протравливании и адсорбции различных веществ, приводящих к увеличению толщины жгутиков (например, после серебрения).
Ворсинки, или пили (фимбрии) - нитевидные образования, более тонкие и короткие (3-10 нм * 0,3-10 мкм), чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина. Известно несколько типов пилей. Пили общего типа отвечают за прикрепления к субстрату, питание и водно-солевой обмен. Они многочисленны - несколько сотен на клетку. Половые пили (1-3 на клетку) создают контакт между клетками, осуществляя между ними передачу генетической информации путем конъюгации (см. главу 5). Особый интерес представляют пили IV типа, у которых концы обладают гидрофобностью, в результате чего они закручиваются, эти пили называют еще кудряшками. Располага-
ются они по полюсам клетки. Эти пили встречаются у патогенных бактерий. Они обладают антигенными свойствами, осуществляют контакт бактерии с клеткой-хозяином, участвуют в образовании биопленки (см. главу 3). Многие пили являются рецепторами для бактериофагов.
Споры - своеобразная форма покоящихся бактерий с грамположительным типом строения клеточной стенки. Спорообразующие бактерии рода Bacillus, у которых размер споры не превышает диаметр клетки, называются бациллами. Спорообразующие бактерии, у которых размер споры превышает диаметр клетки, отчего они принимают форму веретена, называются клостридиями, например бактерии рода Clostridium (от лат.Clostridium - веретено). Споры кислотоустойчивы, поэтому окрашиваются по методу Ауески или по методу Циля-Нельсена в красный, а вегетативная клетка - в синий цвет.
Спорообразование, форма и расположение спор в клетке (вегетативной) являются видовым свойством бактерий, что позволяет отличать их друг от друга. Форма спор бывает овальной и шаровидной, расположение в клетке - терминальное, т.е. на конце палочки (у возбудителя столбняка), субтерминальное - ближе к концу палочки (у возбудителей ботулизма, газовой гангрены) и центральное (у сибиреязвенной бациллы).
Процесс спорообразования (споруляция) проходит ряд стадий, в течение которых часть цитоплазмы и хромосома бактериальной вегетативной клетки отделяются, окружаясь врастающей цитоплазматической мембраной, - образуется проспора.
В протопласте проспоры находятся нуклеоид, белоксинтезирующая система и система получения энергии, основанная на гликолизе. Цитохромы отсутствуют даже у аэробов. Не содержится АТФ, энергия для прорастания сохраняется в форме 3-глицеринфосфата.
Проспору окружают две цитоплазматические мембраны. Слой, окружающий внутреннюю мембрану споры, называется стенкой споры, он состоит из пептидогликана и является главным источником клеточной стенки при прорастании споры.
Между наружной мембраной и стенкой споры формируется толстый слой, состоящий из пептидогликана, имеющего много сшивок, - кортекс.
Кнаружи от внешней цитоплазматической мембраны расположена оболочка споры, состоящая из кератиноподобных белков, со-
держащих множественные внутримолекулярные дисульфидные связи. Эта оболочка обеспечивает резистентность к химическим агентам. Споры некоторых бактерий имеют дополнительный покров -экзоспориум липопротеиновой природы. Таким образом формируется многослойная плохо проницаемая оболочка.
Спорообразование сопровождается интенсивным потреблением проспорой, а затем и формирующейся оболочкой споры дипиколиновой кислоты и ионов кальция. Спора приобретает термоустойчивость, которую связывают с наличием в ней дипиколината кальция.
Спора долго может сохраняться из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизма. В почве, например, возбудители сибирской язвы и столбняка могут сохраняться десятки лет.
В благоприятных условиях споры прорастают, проходя три последовательные стадии: активации, инициации, вырастания. При этом из одной споры образуется одна бактерия. Активация - это готовность к прорастанию. При температуре 60-80 °С спора активируется для прорастания. Инициация прорастания длится несколько минут. Стадия вырастания характеризуется быстрым ростом, сопровождающимся разрушением оболочки и выходом проростка.
Пили типа 1
Пили типа 1 прочно связаны с клеткой, и для того, чтобы отсоединить их от неё, нужны значительные усилия, большие, нежели для удаления жгутиков или половых пилей. Пили данного типа также устойчивы и к химическим воздействиям - сохраняются в 6 М мочевине , 1 М NаОН , устойчивы к додецилсульфату натрия и трипсину . Эти пили разрушаются только при кипячении в растворе с низким значением , что вызывает необратимую денатурацию белка . Белок , образующий пили общего типа 1, имеет молекулярную массу 17 кДа .
Пили типа 1 располагаются перитрихиально, то есть по всей поверхности бактерии. У одной клетки может быть 50-400 пилей длиной до 1,5 мкм. Диаметр этих пилей около 7 нм, а отверстия - 2,0-2,5 нм.
Формирование пилей общего типа 1 определяется генами , расположенными в хромосоме . Их активность подвержена фазовым вариациям, то есть ген может быть активен либо нет. Обычно в культуре присутствуют как клетки, имеющие много пилей общего типа 1, так и лишенные их. Клетки, находящиеся в той или иной фазе, могут быть легко выведены. Размножению клеток, лишенных пилей, способствует выращивание культуры на агаре , тогда как клетки с пилями получают преимущество при выращивании культуры в жидкой среде без аэрации. При этом они образуют пленку. Пили типа 1 придают бактериям гидрофобность , снижают их электрофоретическую подвижность. Они вызывают агглютинацию эритроцитов за счет того, что такие бактерии приклеиваются к эритроцитам (так же, как к другим клеткам животных), а также к клеткам растений и грибов , к неорганическим частицам. В присутствии маннозы нарушается гемагглютинация и прикрепление бактерий к животным клеткам вообще, поскольку пили типа 1 прикрепляются к поверхностным рецепторам , содержащим маннозу . В присутствии маннозы соответствующие участки пилей заняты её молекулами . Адгезивность пилей зависит также от гидрофобности образующего их белка пилина. С маннозными рецепторами реагируют участки пилей, расположенные по всей их поверхности, тогда как за гидрофобные взаимодействия ответственны окончания пилей.
Пили типа 2
Пили типа 2 сходны с пилями 1-го типа, но не вызывают агглютинации эритроцитов, не способствуют образованию бактериями пленки в жидкой среде. Антигенно они близки к пилям 1-го типа и, по-видимому, представляют собой их мутантную форму. Описан и еще ряд вариантов пилей, близких к пилям 1-го типа. Связи пилей общего типа 1 с патогенностью у штаммов Е. coli не удается обнаружить. У энтеропатогенных штаммов обычно образуются другие пили, кодируемые плазмидными генами. Известно несколько типов таких пилей, причем обнаруживается связь типа пилей со специфичностью бактерий в отношении тех или иных животных.
Другие типы пилей
Пили, известные как антигены К88 и К99, тоньше и лабильнее пилей 1-го типа. Они вызывают гемагглютинацию, устойчивую к маннозе, и способствуют прикреплению бактерий к клеткам кишечного эпителия животных, но не человека . Пили 987Р определяют способность Е. соli прикрепляться к эпителию тонкого кишечника новорожденных свиней ; морфологически они похожи на пили 1-го типа. Пили, определяемые генетическим фактором СFА/1, вызывают агглютинацию человеческих эритроцитов и найдены у патогенных для человека штаммов. Молекулярная масса белков пилинов, кодируемых плазмидными генами, 14,5-26,2 кДа. У энтеропатогенных штаммов Е. соli пили являются одним из факторов патогенности, обеспечивающим им возможность прикрепления к клеткам кишечного эпителия. Колонизация бактериями эпителия способствует эффективному взаимодействию выделяемого ими энтеротоксина с клетками эпителия. В результате происходит нарушение водного обмена ткани, что клинически проявляется как диарея . При этом бактерии энергично размножаются в тонком кишечнике , а затем в большом количестве выносятся в окружающую среду, что способствует их распространению.
Половые пили
Половые пили Е. соli образуются у клеток донорских штаммов, отличающихся от изогенных реципиентных наличием у клеток особого генетического детерминанта - полового фактора, или фактора трансмиссивности, который либо является автономным репликоном (F-фактор), либо входит в состав автономного репликона, либо интегрирован с бактериальной хромосомой. Фактор трансмиссивности находится в составе плазмид - факторов множественной устойчивости к антибиотикам (R-факторы), факторов колициногенности и ряда других плазмид. Половые пили отличаются от пилей общего типа по строению и антигенной специфичности, пили, кодируемые различными генетическими детерминантами, также различны.
Половые F-пили, определяемые F-факторами, представляют собой белковые цилиндры, перпендикулярные поверхности клетки, толщиной 8,5-9,5 нм и длиной до 1,1 мкм. Они легко могут быть отделены от клетки при встряхивании бактериальной массы. F-пили образованы белком с молекулярной массой 11,8 кДа. В составе F-пилина отсутствуют пролин , цистеин , гистидин , аргинин . К молекуле пилина присоединены две фосфатные группы и остаток D-глюкозы , связанные с белком ковалентными связями . Пилин содержит довольно много кислых и гидрофобных аминокислот . Он синтезируется на рибосомах , связанных с цитоплазматической мембраной и в цитоплазме не обнаруживается. Пул пилина, видимо, накапливается в цитоплазматической мембране. Его молекулы в процессе синтеза содержат дополнительную сигнальную последовательность аминокислот , отщепляющуюся при транспорте через мембрану. F-пили легко диссоциируют в растворах додецилсульфата натрия и разрушаются органическими растворителями, что связано с гидрофобностью пилина. Бактерии, имеющие F-пили, приобретают новый антиген, у них изменяется поверхностный заряд. Бактерии с F-пилями малоподвижны, проявляют тенденцию к автоагглютинации, например, при понижении значения рН среды. Это также происходит за счет богатства пилина кислыми и гидрофобными аминокислотами. F-фактор интересен еще и потому, что иногда (примерно в 1 случае из 100000) он встраивается в молекулу основной ДНК клетки-хозяина. Тогда при конъюгации переносится не только F-фактор, но, также и остальная ДНК. Этот процесс занимает примерно 90 минут, но клетки могут расходиться и раньше, до полного обмена ДНК. Такие штаммы постоянно передают всю или большую часть своей ДНК другим клеткам. Эти штаммы называются Hrf-штаммами (High frequency recombination), потому что донорная ДНК таких штаммов рекомбинирует с ДНК реципиента.
Для образования F-пилей необходима активность, по крайней мере, 13 генов. Сборка трубочек пилей происходит на цитоплазматической мембране в местах её контакта с внешней мембраной. Трубочка пили проходит через слои муреина и внешнюю мембрану. Для сборки и сохранения пилей необходима энергия . Образованию пилей препятствуют цианид , динитрофенол , азид натрия . Возможно, в процессе сборки происходит фосфорилирование пилина. Обычно клетки с дерепрессированным F-фактором образуют 1-2 пили, а в анаэробных условиях и на богатой среде - до 5 пилей. Причина стимуляции пилеобразования в анаэробных условиях неизвестна. У клеток с оторванными пилями быстро отрастают новые, за 30 секунд пиля достигает 1/2 нормальной длины, а полностью формируется за 4-5 мин. Сформированные пили сохраняются на поверхности клетки 4-5 мин, а затем сбрасываются. Это свидетельствует в пользу точки зрения о том, пили - активные образования. Пили, определяемые фактором Соl I, образованы иным пилином, на них не адсорбируются фаги , специфичные для F-пилей, но имеются специфичные для них фаги. Так называемые мужские фаги адсорбируются на половых пилях, РНК -содержащие фаги - на их боковых поверхностях и нитчатые фаги, содержащие одноцепочечную ДНК, - на кончиках этих пилей. Нитчатый фаг препятствует конъюгации.
При конъюгации к реципиентной клетке присоединяется конец половой пили, при этом рецептором служит белок внешней мембраны реципиентной клетки. Сначала этот контакт не очень прочный и легко может быть нарушен при гидродинамических воздействиях. При этом пары распадаются при множественном заражении РНК-содержащими фагами или в присутствии ионов Zn 2+ . Через несколько минут контакт становится более прочным, происходит сближение клеток и образование между ними цитоплазматического мостика. Имеются данные, свидетельствующие о том, что передача ДНК может происходить и без образования цитоплазматического мостика, а непосредственно через отверстие в пиле. Инактивация пилей антисывороткой и любые повреждающие их воздействия приводят к нарушению процесса конъюгации, в то время как нарушение целостности внешней мембраны или муреинового слоя до некоторого предела влияют на донорские свойства клетки, имеющей пили. После установления контакта с реципиентной клеткой черв пилю в донорскую клетку передается сигнал, вызывающий начало конъюгационного синтеза ДНК. Механизм работы половых пилей еще окончательно не установлен. Ряд наблюдений свидетельствует в пользу модели, предполагающей активную функцию пилей. Согласно этой точке зрения после установления контакта с клеткой реципиента или с вирусом пиля сокращается или втягивается в клетку. Эта модель подтверждается как косвенными, так и прямыми наблюдениями. На электронно-микроскопических препаратах можно проследить, как после адсорбции нитчатого мужского фага на их кончиках пили укорачиваются, а затем нити фага оказываются на поверхности клетки. Сокращение пилей вызыват KCN или арсенат . После воздействия этими ингибиторами пили не обнаруживаются ни на поверхности клеток, ни в окружающей среде, но можно наблюдать адсорбцию на поверхности клеток мужских фагов и антител , специфичных к концам пилей, то есть их кончики, видимо, продолжают выступать над поверхностью клетки. При фаговой инфекции в дальнейшем происходит растворение белковой оболочки нитчатого фага в цитоплазматической мембране бактерии и освобождение его ДНК в цитоплазму. При инфицировании РНК-содержащими мужскими фагами сначала образуется комплекс фаговой РНК с пилином, а фаговый капсид освобождается в среду.
Обычно синтез пилина находится под контролем цитоплазматических репрессоров. В некоторых случаях удается наблюдать определенные закономерности в регуляции образования пилей. Так, в случае Соl I-фактора каждая клетка, получившая при конъюгации плазмиду Соl I, образует пили, их активное образование происходит у клеток 4-8 последующих генераций. Однако затем только единичные клетки в популяции образуют пили, поскольку у большинства бактерий синтез пилина репрессирован. Подобная репрессия, как считают, имеет приспособительное значение, поскольку клетки без пилей не чувствительны к мужским бактериофагам, которые могли бы уничтожить всю популяцию. Единичные клетки с пилями способны обеспечить конъюгацию. При контакте таких клеток с популяциями реципиентных бактерий начинается лавинообразное распространение плазмиды, поскольку образование пилей сначала не репрессировано.
Половые пили обычно образуют только активно растущие клетки, клетки из культуры, находящейся в стационарной фазе роста, обычно лишены пилей и являются плохими донорами.
Как уже было отмечено, существует много более или менее различающихся плазмид, способных определять образование половых пилей, которые также несколько различаются. Рецепторы на поверхности реципиентных клеток обладают разной степенью сродства к разным пилям, что может сильно влиять на эффективность конъюгации бактерий.
Пили, подобные пилям E. coli , образуют и другие представители Enterobacteriaceae . Половые пили имеют Vibrio , Pasteurella , Aeromonas , Pseudomonas .
ИСТОРИЯ И ОБЩАЯ МИКРОБИОЛОГИЯ
1.К исследованиям Луи Пастера относится все перечисленное, кроме:
Г. Открытие возбудителя сибирской язвы.
2. К работам Р. Коха относится все перечисленное, кроме:
Е. Получение вакцины против сибирской язвы.
3. К заслугам И.И. Мечникова относится все перечисленное, кроме:
Е. Создание учения о роли антител в появлении иммунитета.
4. К работам Д.И. Ивановского относится все перечисленное, кроме:
Г. Показал, что вирусы видны в электронный микроскоп.
5. К палочковидным формам относятся все, кроме:
A. Стафилококки.
6. Палочковидные бактерии различаются по всем перечисленным признакам, кроме:
B. Количеству завитков.
7. Фиксация мазков из культур микробов проводится:
A. В пламени спиртовки.
8. Цель фиксации мазков:
A. Прикрепление бактерий к стеклу.
9. Простые методы окраски позволяют выявить все признаки, кроме:
B. Особенностей строения и химического состава некоторых структур клетки.
10. Для простых методов окрашивания используют:
A. Фуксин Пфейффера водный.
11. Какая из перечисленных структур бактерий определяет их способность прикрепляться к поверхности клеток: .
B. Микроворсинки (пили).
12. Для структуры клеточной стенки бактерий характерны все нижеуказанные свойства, кроме
Г. Отвечает за процесс дыхания бактерий.
13. В цитоплазматической мембране происходят все процессы, кроме:
B. Движения бактерий.
Б. Плазмид.
15. К свойствам спор бактерий относятся все, кроме:
Г. Являются формой размножения бактерий.
16. К свойствам капсул бактерий относятся все, кроме:
A. Капсулы хорошо воспринимают красители.
17. Для окрашивания по методу Грама требуется все, кроме:
B. Ацетон.
18. Для окраски мазка по методу Циля-Нильсена требуется все, кроме:
Г. Этиловый спирт.
19. Для окраски мазка по методу Гинса-Бурри требуется все, кроме:
A. Фуксин Пфейффера (водный).
20. Стерилизация - это:
Б. Обеспложивание - полное уничтожение всех микроорганизмов и их спор.
21. Обеззараживание объектов внешней среды - это:
B. Дезинфекция.
22. Дробной стерилизацией называется:
Б. Тиндализация.
23. Искусственные питательные среды классифицируют по всем ниже перечисленным признакам, кроме:
24. К искусственным питательным средам предъявляются все ниже перечисленные требования, кроме:
Ж. Наличие спирта.
25. При бактериоскопическом исследовании материала можно определить все перечисленное, кроме:
Г. Антигенной структуры бактерий.
26. Бактериологическое исследование материала проводят с целью:
B. Определения вида бактерий.
27. К видовым (таксономическим) свойствам бактерий не относятся:
Г. Способность окрашиваться фуксином Пфейффера.
28. Бактериофаг характеризуется всеми перечисленными признаками, кроме:
A. Клеточной организацией.
29. К осложнениям антибиотикотерапии можно отнести все перечисленное, кроме:
Б. Лизогенная конверсия.
30. Продуцентами антибиотиков не являются:
Б. Бактериофаги.
31. В классификации бактерий по типу дыхания нельзя указать следующую группу: .
Б. Аутотрофы.
32. В процессе дыхания у анаэробов принимают участие:
Г. Дегидраза.
ИНФЕКЦИЯ И ИММУНИТЕТ
33. Периоды развития инфекционного процесса все, кроме:
Г. Суперинфекции.
34. Как называются инфекционные болезни, источником которых являются объекты окружающей среды:
Д. Сапронозы.
35. Повторные проявления заболевания, вызванные тем же самым возбудителем - это:
А.Рецидив.
36. Продромальный период - это период:
Г. Появления неспецифических симптомов инфекционной болезни.
37. Для септикопиемии характерно все, кроме:
Д. Отсутствия в крови патогенных микроорганизмов.
38. Характерное свойство эндотоксинов:
Б. Находятся в клеточной стенке грамотрицательных бактерий.
39. Для бактериальных экзотоксинов характерно все, кроме:
B. Липополисахаридная природа.
40. Адгезивная способность бактерий обусловлена:
Г. Наличием пилей.
41. К факторам патогенности бактерий относятся все, кроме:
Д. Отсутствия оформленного ядра.
42. Фактором патогенности с инвазивной функцией является:
B. Мечников И.И.
45. Активным иммунитетом является:
A. Поствакцинальный. Б. Постинфекционный B. Формирующийся при введении анатоксина.
Г. Возникающий при ревакцинации.
Д. Все перечисленное.
46. Какой вид иммунитета является пассивным:
А. После введения иммунных сывороток.
47. Центральным органом иммунитета является:
В. Костный мозг.
48. К иммунокомпетентным клеткам относятся все, кроме:
Д. Эритроцитов.
49. Полноценные антигены обладают:
A. Высокой молекулярной массой. Б. Иммуногенностью.B.Специфичностью.Г.Чужеродностью.
Д. Всем вышеперечисленным.
50. Что такое гаптен?
Д. Неполный антиген.
51. Какое из перечисленных химических веществ является полноценным антигеном?
52. Чем определяется специфичность белкового антигена?
Г. Качественным составом и последовательностью аминокислот в эпитопе.
53. Специфичность антител определяется:
В. Пространственной конфигурацией активного центра.
54. К факторам неспецифической резистентности относятся:
А. Фагоцитоз. Б. Лизоцим. В. Нормальная микрофлора. Г. Интерферон.
Д. Все вышеперечисленное.
55. Для иммуноглубулинов класса М характерно:
A. Способность связывать комплемент. Б. Появляется первым в иммунном ответе на инфекцию.
B. Состоит из пяти субъединиц. Г. Является самым тяжелым из всех классов иммуноглобулинов.
Д. Все вышеперечисленное.
56. Отличительной особенностью иммуноглобулинов класса Е является:
B. Цитофильность.
57. Иммуноглобулины класса А:
A. Обеспечивают местный иммунитет.
58. Наличие полного гемолиза при постановке РСК расценивается как:
B. Отрицательный результат реакции.
59. Образование в титрационной панели аморфного осадка в виде «перевернутого зонтика» можно расценивать как:
A. Положительный результат при постановке РНГА.
60. Для выявления антител в сыворотках больных людей используют:
61. «Меченые» (коньюгированные) диагностические сыворотки используют для выявления микробных антигенов в реакции:
Д. Иммунофлюоресценции
62. Для специфической профилактики заболеваний, вызванных токсинообразующими бактериями, используют: В. Анатоксины
СТАФИЛОКОККИ
63. Стафилококки окрашиваются по Граму:
А. Положительно.
64. Расположение стафилококков в мазке:
B. Группами («виноградная гроздь»).
65. Питательная среда, используемая для культивирования стафилококков:
66. Какое из нижеперечисленных свойств стафилококков дает основание считать их вирулентными:
B. Коагулазная активность.
67. Основной метод диагностики стафилококковых инфекций:
Б. Бактериологический.
68. Обязательно ли изучение антибиотикочувствительности выделенных культур патогенных стафилококков:
СТРЕПТОКОККИ
69. Питательная среда для культивирования стрептококков:
Б. Кровяной агар.
70. Характер гемолиза подозрительных колоний пиогенных стрептококков:
A. Бета-гемолиз.
71. Принцип классификации стрептококков:
Б. По антигенной структуре.
72. Постинфекционный иммунитет после перенесенной стрептококковой ангины:
Б. Непродолжительный.
МЕНИНГОКОККИ
73. Расположение в мазке менингококков:
B. Парами.
74. Что считают главным фактором вирулентности менингококка:
Г. Способность к выживанию внутри клетки.
75. Питательные среды для культивирования менингококков:
B. Сывороточный агар.
76. Вакцина, используемая для профилактики менингококковых инфекций:
Б. Химическая.
77. Окраска менингококков по Граму:
Б. Отрицательная.
78. Какой из сахаров ферментируют гонококки:
А. Глюкоза.
79. Постинфекционный иммунитет при гонорее:
Б. Не выражен.
80. Гоновакцина применяется для:
А. Провокации.
БРУЦЕЛЛЕЗ
81. Морфология бруцелл:
Б. Короткие грамотрицательные палочки.
82. Какие животные являются природным резервуаром В. abortus :
B. Крупный рогатый скот.
83. Основной метод, используемый для диагностики бруцеллеза:
Б. Серодиагностика.
84. Для профилактики бруцеллеза используют:
A. Живую вакцину.
СИБИРСКАЯ ЯЗВА
85. Морфология В. anthracis :
Б. Стрептобациллы.
86. Продукция сибиреязвенного экзотоксина:
Б. Не продуцируют.
87. В реакции Асколи исследуют:
В. Материал от погибших животных
88. Особенность сибиреязвенной вакцины:
A. Бескапсульные авирулентные микроорганизмы.
89. При микроскопическом исследовании материала обращают внимание на следующие особенности возбудителя чумы:
Б. Овоидная форма с биполярным окрашиванием.
90. Характер колоний Y . pestis :
B. Колонии R-формы («смятый кружевной платочек»).
91. Степень выраженности постинфекционного противочумного иммунитета:
B. Напряженный стойкий иммунитет.
92. К какой группе биологически опасных агентов относится возбудитель чумы:
A. Первая.
ТУЛЯРЕМИЯ
93. Основным резервуаром возбудителя в природе являются:
B. Грызуны.
94. Для получения первых генераций возбудителя туляремии используют:
Б. Чувствительных лабораторных животных.
95. Аллергическая проба с тулярином выявляет:
96. Для профилактики туляремии используют:
В. Живую вакцину
КИШЕЧНЫЕ ИНФЕКЦИИ
97. Энтеробактерии окрашиваются по Граму:
Б. Отрицательно.
98. У энтеробактерии тип дыхания:
B. Факультативноанаэробный.
99. Для всех энтеробактерий характерна утилизация:
A. Глюкозы.
100. О-антиген энтеробактерии представляет собой:
Г. Липополисахаридопротеиновый комплекс.
101. Иммуногенность энтеробактерий обусловливает:
Б. Полисахаридная часть.
102. У новорожденных наиболее частым возбудителем ОКИ являются:
B. Энтеропатогенные эшерихии.
103. Для первичного выделения эшерихий не применяют:
B. Кровяной агар.
104. Дифференциация внутри вида E . coli основана главным образом на изучении:
Г. Антигенной структуры.
105. Ha пластинчатых средах 2 типа (S и R ) колоний образуют:
106. Самый активный по биохимическим свойствам вид шигелл:
107. Для рода шигелл общим и стабильным является:
A. Отсутствие подвижности
108. Методом ранней диагностики брюшного тифа является:
A. Выделение гемокультуры.
109. Кровь от больного с подозрением на брюшной тиф следует сеять:
Б. На среду Раппопорт.
110. Классификация сальмонелл основана на различиях в:
A. Антигенной структуре.
111. Серологический метод диагностики брюшного тифа и паратифов был разработан:
A. Видалем.
112. Наиболее часто в настоящее время выделяются от больных сальмонеллы вида:
A. S. enteritidis.
113. При брюшном тифе для серологической диагностики предпочтительной является:
114. В качестве среды обогащения при сальмонеллезе используют все, кроме:
Г. Солевой бульон.
115. При диагностике кишечного сальмонеллеза материалом исследования служит все, кроме:
Г. Мокрота.
116. К антропонозным заболеваниям не относится:
Б. Сальмонеллез.
117. Разложение углеводов до кислоты без газа характерно для сальмонелл:
118. Главными тестами в идентификации выделенных культур рода шигелла являются:
A. Биохимическая активность.
Б. Тест антигенной структуры.
B. Лизабельность специфическим бактериофагом.
Г. Все перечисленное.
119. Основным признаком дифференциации биоваров возбудителей холеры является:
Г. Чувствительность к специфическим бактериофагам.
120. Главный фактор патогенности возбудителя холеры:
B. Энтеротоксин.
121. Основным признаком, идентифицирующим вид возбудителя холеры, является:
В. Антигенная структура.
122. Возбудителями холеры не является:
A. V. parahaemoliticus.
123. Дисбактериозом кишечника называют:
Б. Количественные и качественные изменения собственной микрофлоры кишечника.
124. Основные функции нормальной микрофлоры макроорганизма:
Е. Все перечисленное.
125. В кишечнике практически здоровых людей должны преобладать:
A. Анаэробы.
126. Обнаружение бифидобактерий производится на:
Г. Среде Блаурока.
127. При дисбиозе в составе эшерихий возможны следующие изменения:
A. Увеличение общего количества эшерихий.
Б. Замена полноценных в ферментативном отношении эшерихий на эшерихии со сниженной ферментативной активностью.
B. Возрастание количества эшерихий с гемолитической активностью.
Г. Снижение общего количества эшерихий.
Д. Все перечисленное.
128. При обследовании на дисбиоз первичный посев производится на:
В. Селективные среды количественным методом.
129. Для пищевых отравлений характерно:
A. Острое внезапное начало.
Б. Одновременность заболевания у группы лиц.
B. Связь заболевания с употреблением какого-то одного продукта или блюда.
Г. Острое короткое течение заболевания.
Д. Все перечисленное.
130. Для человека патогенны:
B. Типы А, В, Е.
131. Для С1. botulinum характерно все, кроме:
А. Принадлежности к Грам (-) флоре.
132. Для стафилококкового пищевого токсикоза характерно:
A. Накопление в пищевом продукте стафилококкового энтеротоксина.
Б. Накопление в пищевом продукте эндотоксина.
133. Возбудителями пищевых токсикозов являются все, кроме:
Б. Протеи.
134. Отравления «продуктами моря» чаще всего связаны с:
Г. V. parahaemoliticus.
135. По патогенетическому принципу микробные пищевые отравления делятся на:
A. Токсикоинфекции и токсикозы.
136. Вид Cl . botulinum подразделяется на варианты по следующим свойствам:
B. Специфика экзотоксина.
КАПЕЛЬНЫЕ ИНФЕКЦИИ
137. Методы окраски, используемые для изучения включений возбудителя дифтерии:
Б. Нейссера.
138. При идентификации возбудителя дифтерии изучают:
B. Токсигенность.
139. Какая особенность токсигенных штаммов Corynebacterium diptheriae :
Б. Лизогенные бактерии.
140. Какой основной признак лизогенных бактерий:
141. Токсигенность возбудителя дифтерии изучают:
A. В реакции диффузионной преципитации в геле.
142. Типы gravis и mitis можно дифференцировать по следующим свойствам:
A. Культуральным и биохимическим.
143. Противодифтерийная антитоксическая сыворотка используется в основном:
B. Для лечения больных дифтерией.
144. АКДС-вакцина включает:
A. Дифтерийный анатоксин.
145. При подозрении на коклюш для бактериологического исследования берут:
A. Материал из зева.
146. Отличить возбудитель коклюша от паракоклюша можно по:
A. Морфологическим и тинкториальным свойствам.
Б. Антигенной структуре.
147. Диагноз коклюша позволяет поставить:
B. Бактериологический метод.
148. Возбудитель паракоклюша растет на:
Б. Мясо-пептонном агаре.
149. Методы окраски, используемые для определения возбудителя туберкулеза:
B. Циля-Нильсена.
150. С чем связана кислотоустойчивость возбудителя туберкулеза:
B. С высоким содержанием липидов в клеточной стенке.
151. Какая проба позволяет выявить сенсибилизацию макроорганизма при туберкулезе:
152. Вид вакцины, применяемой для профилактики туберкулеза:
СПИРОХЕТЫ
153. Для диагностики первичного сифилиса применяется:
Б. Микроскопия исследуемого материала.
154. Особенности микроскопии бледной трепонемы:
Б. Темнопольная микроскопия нативного препарата.
155. Какие тесты наиболее специфичны для диагностики сифилиса:
В. Реакция иммобилизации трепонем
156. Из какой культуры спирохет получают ультраозвученный трепонемный антиген
Б. Выращенный в тестикулярной ткани кроликов
РИККЕТСИИ
157. Риккетсии Провачека у человека вызывают:
В. Эндемический сыпной тиф
158. Для изучения морфологии риккетсий Провачека применяется следующий метод окраски:
Г. Романовского-Гимза
159. Наиболее эффективный метод культивирования риккетсий Провачека:
Г. В куриных эмбрионах
160. К факторам вирулентности риккетсий Провачека следует отнести
В. Наличие микрокапсулы.
161. Какая вакцина может быть использована для профилактики эпидемического сыпного тифа
Б. Химическая
162. Для дифференцировки эпидемического сыпного тифа и болезни Бриля используют
Г. Определение класса антител в сыворотке больного.
163. Для диагностики Ку-лихорадки применяется следующие методы:
А. Биологический и серологический
164. Для профилактики Ку-лихорадки используется вакцина:
165. Для серодиагностики волынской лихорадки используют следующие реакции:
ХЛАМИДИИ. МИКОПЛАЗМЫ
166. Хламидии поражают следующие клетки:
B. Эпителиальные.
167. Какие Методы диагностики хламидиозов можно использовать:
Б. Выявлении антител в сыворотке крови больных.
168. Колонии микоплазм характеризуются:
Б. Напоминают «яичницу-глазунью».
АНАЭРОБНЫЕ ИНФЕКЦИИ
169. Основным фактором вирулентности возбудителей столбняка и газовой гангрены являются:
Б. Экзотоксин.
170. Для лечения больных столбняком и газовой гангреной используют:
Г. Антитоксическую сыворотку.
171. Для лабораторной диагностики столбняка применяют:
Б. Биологический метод.
172. Фактором вирулентности грамотрицательных НАБ является наличие:
Б. Эндотоксина.
173. Неклостридиальную анаэробную инфекцию (НАИ) вызывают:
Б. Бактероиды.
ВИРУСЫ И ВИРУСНЫЕ ИНФЕКЦИИ
174. Вирусы отличаются от бактерий по:
В. Способу размножения.
175. Величина вирусов определяется с помощью:
Г. Ультрацентрифугирования.
176. Сложно устроенные вирусы имеют дополнительную внешнюю оболочку:
B. Суперкапсид.
177. Вирусы культивируют:
Г. В культурах клеток.
178. Симпласт - это:
B. Гигантская многоядерная клетка.
179. Световой микроскоп применяется при вирусологических исследованиях для оценки:
Г. Цитопатического действия.
180. Для выделения культуры вируса гриппа чаше всего используют:
Г. Культуру клеток из почек эмбриона человека.
181. Тип гемагглютинина вирусов гриппа можно определить в:
182. Для профилактики гриппа в настоящее время используют:
A. Живую вакцину.
Б. Инактивированную цельновирионную вакцину.
B. Иммуноглобулин.
Г. Субвирионную вакцину.
Д. Все перечисленное
183. Возбудитель парагриппа:
Г. Обладает гемадсорбирующими свойствами.
184. При парагриппозной инфекции реакция гемадсорбции применяется для:
Г. Идентификации вирусов парагриппа.
185. Вирус кори можно культивировать:
B. В культуре клеток.
186. Для серопрофилактики кори используют:
Г. Специфический иммуноглобулин.
187. Респираторно-синцитиальный вирус относится к семейству:
Б. Парамиксовирусов.
188. Местный противовирусный иммунитет при респираторно-синцитиальной вирусной инфекции обусловлен:
189. Материалом для выделения культуры аденовирусов служит:
Б. Фекалии.
B. Отделяемое носоглотки.
Г. Отделяемое конъюктивы.
Д. Все перечисленное.
190. Для аденовирусов характерно :
Б. Наличие ДНК.
191. Заражение полиомиелитом может происходить:
A. Воздушно-капельным путем.
192. Для лабораторной диагностики полиомиелита используется:
Г. Реакция тканевой нейтрализации.
193. Вакцина против полиомиелита относится к типу:
B. Живых вакцин.
194. Вирусы Коксаки:
A. Относятся к пикорнавирусам.
195. Исследуемым материалом в раннем периоде Коксаки-вирусной инфекции является:
A. Смыв из зева.
196. Для идентификации ЕСНО-вирусов с помощью реакции тканевой нейтрализации необходимо иметь:
Д. Диагностические сыворотки против ЕСНО-вирусов.
197. Ведущим механизмом передачи гепатита А является:
A. Фекально-оральный.
198. Для серодиагностики гепатита А используют реакцию: .
199. Геном вируса гепатита В представлен:
Б. Двунитчатой кольцевой ДНК с однонитчатым участком.
200. В сыворотке крови больного гепатитом В в начале болезни можно обнаружить:
Б. HBs-антиген.
201. Открывателем клещевого энцефалита является:
Б. Зильбер Л.А.
202. В диагностике бешенства применяют:
Г. Обнаружение телец Бабеша-Негри.
203. Для экстренной специфической профилактики бешенства при укусах опасной локализации применяют:
Б. Инактивированную культуральную вакцину.
204. Один из путей передачи клещевого энцефалита:
Б. Алиментарный.
205. Переносчиком и резервуаром вируса клещевого энцефалита одновременно могут являться:
206. В ранней лабораторной диагностике клещевого энцефалита используется:
Г. Определение антител класса М в сыворотке больного при помощи ИФА.
207. К ВИЧ наиболее чувствительны следующие клетки:
Г. Т- хелперы.
208. Поверхностным антигеном ВИЧ – вируса является белок:
209. Способность к интеграции в геном клетки связана у ВИЧ с:
Г. Наличием обратной транскриптазы.
210. Особенностью герпесвирусов является:
Б. Способность к персистенции.
211. Вирус ветряной оспы вызывает:
B. Опоясывающий герпес.
212. Для подтверждения активной герпетической инфекции проводят:
B. Поиск Ig M при помощи ИФА.
213. Для специфической терапии герпетической инфекции применяют: